
Recomendados
Más contenido relacionado
La actualidad más candente
La actualidad más candente (20)
Destacado
Similar a Cell adhesion in plants and animals
Similar a Cell adhesion in plants and animals (20)
Último
Último (20)
Cell adhesion in plants and animals
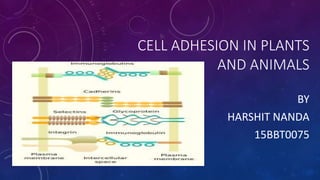
- 1. CELL ADHESION IN PLANTS AND ANIMALS BY HARSHIT NANDA 15BBT0075
- 2. METHODS OF CELL ADHESION Cells adhere by cell junctions, cell adhesion molecules, or substrate adhesion molecules
- 3. TYPES OF CELL ATACHMENTS • Tight junctions are regions where membranes of adjacent cells actually fuse. • They encircle the whole cell and provide a barrier for leakage between cells. • Desmosomes are spot rivets that weld cells together. • A cytoplasmic plaque is connected by inter cellular protein filaments, and the plaques are connected to the intermediate filaments within the cytoplasm • Gap junctions are small channels between cells that allow for intercellular communication
- 4. CELL ADHESION MOLECULES Cell adhesion molecules (CAMs) are glycoproteins with 3 major domains: The extracellular domain allows one CAM to bind to another on an adjacent cell. The binding can be to the same type of cell (homotypic) or to a different cell type (heterotypic). The transmembrane domain links the CAM to the plasma membrane through hydrophobic forces. The cytoplasmic domain is directly connected to the cytoskeleton by linker proteins. This anchoring is important to prevent lateral diffusion of adhesion molecules in the membrane.
- 5. Cadherins mediate calcium- dependent cell adhesion Cadherins are the most prevalent CAMs in vertebrates. They are rapidly degraded by proteases in the absence of Ca++. There are 4 major types: E cadherins in epithelial cells P cadherins in placenta N cadherins in neural tissue L cadherins in liver Each associates with its own type.
- 6. SUBSTRATE ADHESION MOLECULES (SAMS) • Spaces between cells are filled with ECM that consists of: • 1. Amorphous ground substance: a gel-like material that absorbs water. • 2. Meshwork of fibers that reinforce the ground substance. • The ECM influences cell migration, cell shape, cell gene expression, and cell differentiation.
- 7. GLYCOSAMINOGLYCANS AND PROTEOGLYCANS FORM THE AMORPHOUS GROUND SUBSTANCE • Glycosaminoglycan: long unbranched polysaccharide chains composed of repeating units of disaccharides. One sugar is an amino sugar (n-acetyl glucosamine) and the other is a uronic acid • Proteoglycans: glycosaminoglycans are covalently linked to core proteins The core proteins have have many side chains of glycosaminoglycans. They attract Na+ and water and expand to form gels that occupy space between cells.
- 8. INTEGRINS MEDIATE ADHESION TO ECM • Integrin are a family of transmembrane glycoproteins that are composed of 2 chains, a and b. There are 40 different types of a chains and 8 types of b chains that can combine to form a large number of different integrin molecules. • The a chain has binding sites for Ca++ and Mg++ which are needed for integrins to adhere. The 2 subunits form the site that binds to the RGD domain on ECM.
- 9. Two main ways in which animal cells are bound together: in connective tissue, the main stress-bearing component is the extracellular matrix. In epithelial tissue, it is the cytoskeletons of the cells themselves linked from cell to cell by anchoring junctions. Cell- matrix attachments bond epithelial tissue to the connective tissue beneath it.
- 10. PLANTS CELL ADHESION The adhesive mechanism at the middle lamella that maintains cell to cell links until separation is not known. A calcium-dependent mechanism involving cross links of homogalacturonan regions of pectic polysaccharides is probably the most widely considered possibility , although this does not seem to have been rigorouslytested. The main evidence for a role for pectins is derived from observations of the action of pectinases and calcium chelators resulting in cell separation in certain tissues. However, the ability of these treatments to result in cell separation varies between tissues
- 11. PLANT TISSUES • The overall structural organization of plants is generally simpler than that of animals. For instance, plants have only four broad types of cells, which in mature plants form four basic classes of tissue: dermal tissue interacts with the environment; vascular tissue transports water and dissolved substances (e.g., sugars, ions); space filling ground tissue constitutes the major sites of metabolism; and sporogenous tissue forms the reproductive organs • Plant tissues are organized into just four main organ systems: stems have support and transport functions; roots provide anchorage and absorb and store nutrients; leaves are the sites of photosynthesis; and flowers enclose the reproductive structures. Thus at the cell, tissue, and organ levels, plants are generally less complex than most animals
- 12. • Unlike animals, plants do not replace or repair old or damaged cells or tissues; they simply grow new organs. • Most importantly in contrast with animals, few cells in plants directly contact one another through molecules incorporated into their plasma membranes • Instead, plant cells are typically surrounded by a rigid cell wall that contacts the cell walls of adjacent cells • Also in contrast with animal cells, a plant cell rarely changes its position in the organism relative to other cells • These features of plants and their organization have determined the distinctive molecular mechanisms by which their cells are incorporated into tissues
- 13. INTERCELLULAR SPACE FORMATION • Intercellular space formation occurs to some extent in all tissues, generally at cell corners. • Their formation usuallyinvolves disruption of a highly localized region of a parent cell wall leading to the connection of a middle lamella between daughter cells with the middle lamella of the parent cell. • It will be of great interest to determine the means of the regulation of this disruption and the relativeinvolvement of the participating cells.
- 14. CELLULAR INTERACTIONS IN PLANTS • Due to the lack of relative cell movement and active adhesion phenomena involving recognition and sorting, the cell signalling requirements for plant cells during development would therefore seem to be distinct fromthose of animal cells. • Cell position with a population of meristematic cells appears to be an important factor in the determination of cell fate . • A component cell of a multicellular structure needs to know if it has neighbours and who they are, i.e. what is its own developmental context. • Whatever the cell signalling mechanisms are, the maintenance of close cell contact observed within meristems and embryos would seem to be a requirement of the setting up of the early cell distinctions. The subsequent development of the cells can be viewed as a gradual relaxation of this tight adherence in a developmentally regulated manner.